| ما هو الحمض الريبي النووي منزوع الأكسجين DNA؟ |
1- النوكليوتيدات هي حجارة بناء الحمض الريبي منزوع الأكسجين DNA
|
يعد جزيء الـ DNA عديد جزيئات متكررة (مكثور) polymer أساسها النوكليوتيد، وهناك أربعة نوكليوتيدات مختلفة مكونة لشريط الـ DNA:
- dATP: ديوكسي ادينوزين ثلاثي الفسفات
- dGTP: ديوكسي غوانين ثلاثي الفسفات
- dTTP: ديوكسي ثيميدين ثلاثي الفسفات
- dCTP: ديوكسي سيتوزين ثلاثي الفسفات
تدعى هذه النوكليوتيدات الأربعة dNTP's (ديوكسي نوكليوزيد ثلاثي الفسفات)، وذلك لأنها تتألف من ثلاثة مكونات رئيسة: أساس آزوتي وجزيء السكر وجزيئة ثلاثي الفسفات. تختلف النوكليوتيدات الأربعة عن بعضها البعض بنوع الأساس الآزوتي الداخل في تركيبها.
|
2- كيف تشكل النوكليوتيدات سلاسل الحمض النووي؟
يتشكل الحمض النووي من مجموعة نوكليوتيدات مرتبطة مع بعضها بروابط هيدروجينة وفق ترتيب وتسلسل معين يشكل الرواميز (الشيفرة) الوراثية التي ستترجم فيما بعد إلى بروتين. |
| الحمض النووي في الخلية |
1- ما هي العضيات Micro-organisms التي تحتوي على الحمض النووي؟
كافـة خلايا العضيات الحقيقيـة النوى Eukariotic تحتوي على الحمض الريبي النووي RNA والحمض الريبي النووي منزوع الأكسجين DNA. تحتـوي النواة ضمن الخلية على معظم الأحماض الريبية النووية ضمن بنية محددة هي ما يسمي بالصبغيات chromosomes. كما تحتوي المتقدرات Mitochondria أيضاً على الحمض الريبي النووي منزوع الأكسجين ويسمى بـ Mitochondrial DNA.
هذا ويعطي الجـدول 1 فكرة عن حجـم
الأحماض الريبية النووية، أو ما يمكن تسميته
بحجم المادة الوراثية عند عدد من الأحياء. ولإعطاء فكرة عن حجم الحمض النووي فإن: كل مليون زوج نوكليوتيدي يشغل حيزاً خطياً قدره 0.34 مم. وإذا أردنا قياس طول الـ DNA عند خلية بشرية واحدة نجد أنها تشكل 204 سم (تحتوي الخلية البشرية على ما مجموعه 6 × 910 زوج نوكليوتيدي).
2- ما هو الهدف من تحديد التسلسل النوكليوتيدي:
إن الهدف من تحديد التسلسـل النوكليوتيدي هو تحديد ترتيب النوكليوتيدات ضمن المورث الهدف والذي يعكس ترتيب الأحماض الأمينية المشكلة للبروتين الناتج عن عملية النسخ والترجمة لرواميز الأحماض النووية الوراثية. تتم أحياناً مقارنة الترتيب النوكليوتيدي الناتج مع الترتيب النوكليوتيدي الأصلي المعروف لكشف الطفرات الوراثية إن وجدت في العينة، وتتم المقارنة آلياً.
3- كيف يمكن الحصول على التسلسل النوكليوتيدي؟
لتحديد التسلسل النوكليوتيدي، لا يتم الانطلاق من كامل المجين genome كما في تفاعل الـ PCR، وإنما يتم البدء من شدفة fragment الـ DNA الهدف أو من المورّث المحمّل على بلازميدةDNA Plasmid.
قبل الحصول على التسلسل النوكليوتيدي يجب أولاً عزل الحمض النووي من الخلية. هناك العديد من الطرق لعزل أو استخلاص الـ DNA لسنا هنا بصدد الحديث عنها. تلي مرحلة عزل الـ DNA مرحلة تضخيم amplification الشدفة الهدف المراد معرفة التسلسل النوكليوتيدي لها، ولهذا القصد يستخدم جهاز تضخيم الـ DNA الذي يعتمد على تفاعل سلسلة البوليميراز Polymerase Chain Reaction (PCR). وتتكـون دورة التضخيم الواحدة من ثلاث خطوات رئيسة، وهي:
1- مرحلة فصل سلسلتي الـ DNA وتُجرى عند درجة حرارة 94oم.
2- مرحلة ارتباط المَشْرَع بالسلسة الهدف وتجرى عند درجة حرارة ما بين 50-65oم وهي تختلف حسب المَشْرَع المستخدم.
3- مرحلة تركيب الشدفة الهدف وتجرى عند درجة حرارة 72oم.
تكرر هذه الدورة مابين 30-40 مرة لنحصل في نهايتها على ما يعادل مليون نسخة
من شدفة الـ DNA الهدف.
4- تحديد التسلسل النوكليوتيدي لشدفة الـ DNA الهدف.
يتم تحديد التسـلسـل النوكليوتيـدي وفق الخطوات التالية:
1- تفاعل تحديد التسلسل النوكليوتيدي:
يتم تحديد التسلسل النوكليوتيدي بدورة عمل تكرر من 30 إلى 40 مرة آلياً، وتتألف كل دورة من المراحل الثلاث التالية (كما في تفاعل الـ PCR):
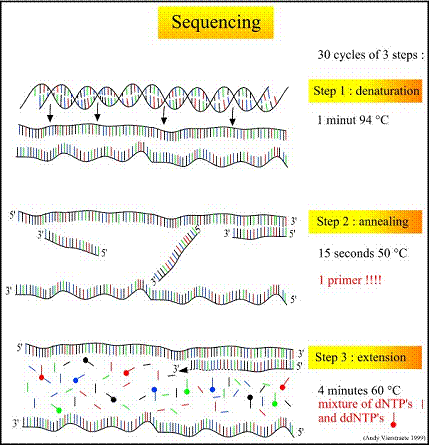
أ. مرحلـة تمسّـخ سلسلتي الـ DNA للشدفة الهدف المضخّمة (Denaturation at 94oC): تتم هذه المرحلة عند درجة حرارة 94oم، لتمسخ الشريط المضاعف لشدفة الـ DNA الهدف. يتعطل في هذه المرحلة عمل جميع الإنزيمات في الوسط (يتوقف على سبيل المثال عمل إنزيم التنسخ من الدورة السابقة).
ب. مرحلة ارتباط المَشْرَع بالسلسة الهدف (التطوِّيع Annealing at 50oC): في تفاعل تحديد التسلسل النوكليوتيدي يستخدم أحد المشرعين المستخدمين في تفاعل الـ PCR الأول المضخم لشدفة الـ DNA الهدف، (يستخدم المَشْرَع الأيسر أو الأيمن)، ولذلك يجرى في هذا التفاعل انتساخ شدفة واحدة فقط من إحدى سلسلتي الـ DNA، بينما في تفاعل الـ PCR العادي يجري استخدام مشرعين (أيمن وأيسر) وبالتالي يجري تضخيم نسخ لكلا السلسلتين معاً. يرتبط المشرع بشدفة محددة من سلسلة الـ DNA المقابلة لها من حيث الأسس الآزوتية، ويتم ذلك الارتباط عبر روابط الفسفور ما بين نوكليوتيدات المشرع ونوكليوتيدات السلسلة الهدف. تجري هذه المرحلة عند درجة حرارة 50oم.
ج. مرحلة الامتداد أو تركيب الشدفة الهدف (Extension at 60oC): وتُجرى عند درجة حرارة 60oم، وتعد هذه الدرجة مثاليةُ لعمل إنزيم البوليميراز polymerase (تكون الدرجة المستخدمة في هذه المرحلة هي 72oم عادةً، ولكن على اعتبار أنه تستخدم هنا نوكليوتيدات ذات نهاية موسومة labeled بمادة متفلورة ddNTP's، فإن درجة الحرارة الفعالة المعتمدة هنا هي 60 بدلاً من 72oم). بعد ارتباط المَشْرَع في المرحلة السابقة، يتم في هذه المرحلة ارتباط النوكليوتيدات (dNTP's أو ddNTP's) إلى السلسة الهدف وتركيب السلسة المكملة ابتداءاً من الطرف 3' باتجاه الطرف 5'، بإضافة كلا النوعين من النوكليوتيدات خلال تركيب السلسلة المكملة، ولكن عندما يضاف نوكليوتيد مفلور dNTP إلى سلسة الهدف تتوقف عملية التركيب لأنه يحتوي على ذرة هيدروجين H على ذرة الكربون الثالثة (يحتوي النوكليوتيد العادي dNTP على وظيفة ماءات OH في هذا الموقع عادة عوضاً عن ذرة الهيدروجين)، وعلى اعتبار أن هذا النوكليوتيد يحوي جزيئاً مفلوراً، فإنه من الممكن تحديد اللون لآخر نوكليوتيد مرتبط مع السلسلة الهدف باستخدام رحلان كهربي خاص وبوجود أشعة الليزر. يوضح الشكل 1 مراحل تحديد التسلسل النوكليوتيدي المختلفة.
يُستخدم المَشْرَع في هذا التفاعل، لذلك يتم تشكل نسخ عديدة متماثلة من إحدى سلسلتي شدفة الـ DNA الهدف، وعلى اعتبار أنه يتم تكرار دورات النسخ، فإنه يحدث تضاعف شدفة الـ DNA الهدف وفق زيادة خطية، أي ستتشكل أعداد كبيرة منها في النهاية. فعلى افتراض أن عملية النسخ بدأت بوجود 1000 نسخة من شدفة الـ DNA الهدف قبل بدء التفاعل، تتشكل بعد دورة واحدة 2000 نسخة (1000 نسخة أصلية و1000 نسخة مكملة وتحوي على النوكليوتيدات المفلورة). بعد دورتين سيكون هناك 2000 نسخة، وبعد ثلاثة دورات 3000 نسخة وهكذا. ويبين الشكل 1 التضاعف الخطي لنسخ شدفة الـ DNA الهدف.
2- فصل الجزيئات المضاعفة بواسطة الرحلان كهربي:
بعد عملية تفاعل تحديد التسلسل النوكليوتيدي، يتم فصل خليط السلاسل الناتج المكون من سلاسل مختلفة ومتدرجة في أطوالها على جهاز رحلان كهربي، وتتكون هلامة الفصل من مادة البولي أكريلاميد، القادرة على فصل شدفتي DNA تختلفان عن بعضهما بنوكليوتيد واحد زيادةً أو نقصاً.
تحتوي شدف الـ DNA على شحنة سالبة بسبب وجود شاردة الفسفور في جزيئات النوكليوتيدات المكونة لها، وبالتالي فهي تهاجر نحو القطب الموجب في الرحلان الكهربي. تهاجر الشدف الأصغر بشكل أسرع من تلك الأكبر، وبالتالي تصطف شدف الـ DNA حسب أطوالها على وسط الفصل.
يبين الشكل 2 انفصال شدف الـ DNA على وسط الفصل ضمن وسط الرحلان الكهربي. |
الجدول 1: حجم الأحماض النووية الريبية.
نوع الحمض الريبي النووي |
الكائن الحي |
عدد الأزواج النوكليوتيدية |
Chromosomal DNA (DNA الصبغي) |
الثدييات |
6 × 910 |
النباتات |
2 × 810 - 2 × 1110 |
الفطر |
2 × 710 - 2 × 810 |
Mitochondrial DNA
(DNA المتقدري) |
الحيوانات |
16 × 310 - 19 × 310 |
النباتات العليا |
150 × 310 - 250 × 410 |
الفطر |
17 × 310 - 78 × 310 |
الطحلب الأخضر |
16 × 310 |
الأوالي الحيوانية
protozoa |
22 × 310 - 40 × 310 |
Chloroplast DNA
(DNA صانعات الكلوروفيل) |
النباتات العليا |
120 × 310 – 200 × 310 |
الطحلب الأخضر |
180 × 310 |
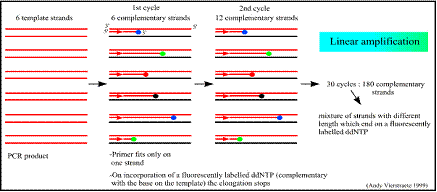
الشكل 1: التضاعف الخطي لنسخ شدفة الـ DNA الهدف. |
- تحديد التسلسل على جهاز الرحلان:
تمر شدف الـDNA المهاجرة والحاوية على النوكليوتيدات المفلورة ضمن هلامة الفصل وفي نهايته الحاوي على نافذة تصدر أشعة ليزرية، عند مرور شدف الـ DNA هذه أمام النافذة الليزرية تصدر Emission ألواناً مختلفة حسب نوع النوكليوتيد، تجمع هذه الإشعاعات اللونية وتوجه نحو كاميرا خاصة وتترجم ضمن برنامج حاسوبي خاص لهذا الهدف. يبين الشكل 3 مبدأ عمل والتقاط الأشعة ضمن الرحلان الكهربي.
كما يبين الشكل 4 كيف تبدو صورة وسط الفصل الناتجة عن عملية الرحلان الكهربي وتحويلها ضمن برنامج حاسوبي خاص إلى ملف صورة.
|
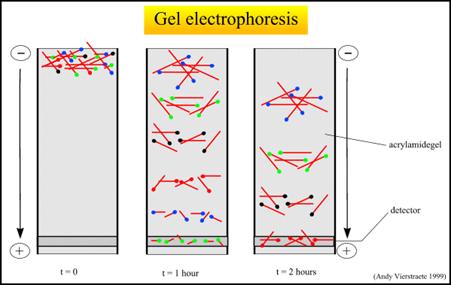
الشكل 2: انفصال شدف الـ DNA على هلامة الفصل ضمن وسط الرحلان الكهربي.
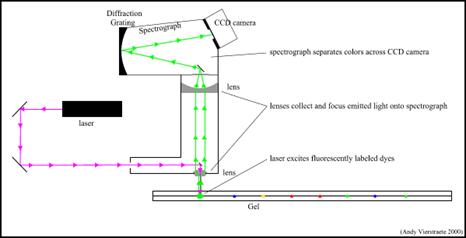
الشكل 3: مبدأ عمل والتقاط الأشعة ضمن الرحلان الكهربي.
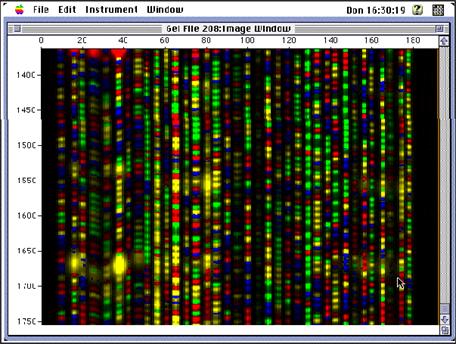
الشكل 4: صورة هلامة الفصل الناتجة عن عملية الرحلان الكهربي وتحويلها ضمن برنامج حاسوبي خاص إلى ملف صورة. |
4- تجميع أجزاء شدف الـ DNA المحددة التسلسل النوكليوتيدي:
يتم تحديد التسلسل النوكليوتيدي للشدفة الهدف من كلا الجهتين (باستخدام المَشْرَع الأيمن والأيسر ضمن تفاعلين منفصلين) ثم يتم التأكد من صحة التسلسل بمقارنة التسلسل الناتج من كلا الجهتين. إن طول الشدفة الهدف التي يمكن تحديد التسلسل النوكليوتيدات لها تترواح من 750 وحتى 800 نوكليوتيداً، وبالتالي إذا كانت الشدفة الهدف أطول من ذلك، يتم تحديد التسلسل النوكليوتيدي لشدف صغيرة ثم تجميع التسلسل النوكليوتيدي لكامل الشدفة الهدف، ولتحقيق ذلك يتم استخدام بادئات مختلف بحيث تغطي الشدفة الهدف.
يبين الشكل 5 تجميع التسلسلات النوكليوتيدية للشدفة الهدف.
|
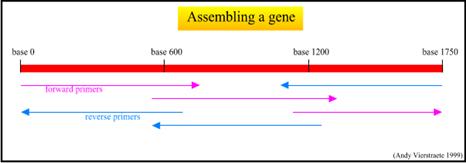
الشكل 5: تجميع التسلسلات النوكليوتيدية للشدفة الهدف. |
| المراجع References |
1-Kocher TD; Thomas WK; Meyer A; Edwards SV; P??bo S; Vilablanca FX. and Wilson AC.
Dynamics of mitochondrial DNA evolution in animals: ampli-fication and sequencing with conserved primers.
Proc. Natl. Acad. Sci. USA, 86: 6196-6200, 1989.
2-Blattner FR; Plunkett III GP; Bloch CA; Perna NT; Burland V; Riley M; Collado-Vides J; Glasner JD; Rode CK; Mayhew GF; Gregor J; Davis NW; Kirkpatrick HA; Goeden MA; Rose DJ; Mau B. and Shao Y.
The complete genome sequence of Escherichia coli K-12.
Science, 277: 1453-1462, 1997.
3-Kawarabayasi Y; Sawada M; Horikawa H; Haikawa Y; Hino Y; Yamamoto S; Sekine M; Baba S; Kosugi H; Hosoyama A; Nagai Y; Sakai M; Ogura K; Otuka R; Nakazawa H; Takamiya M; Ohfuku Y; Funahashi T; Tanaka T; Kudoh Y; Yamazaki J; Kushida N; Oguchi A; Aoki K; Nakamura Y; Robb TF; Horikoshi K; Masuchi Y; Shizuya H. and Kikuchi H.
Complete sequence and gene organization of the genome of a hyper-thermophilic Archae-bacterium, Pyrococcus horikoshii OT3.
DNA Res; 5: 55-76, 1998.
4-Selander RK; Li J; Boyd EF; Wang FS. and Nelson K.
DNA sequence analysis of the genetic structure of populations of Salmonella enterica and Escherichia coli. In F. G. Priest, et al. (ed.), Bacterial diversity and systematics. Plenum Press, New York, 17-49, 1994.
|
| |
