سـتخدمت فئـران ذكور (عدد50 ) من نوع
Sprague-Dawley من مختلف الأعمار، وهي الأكثر توافراً والأسهل استخداماً والتي
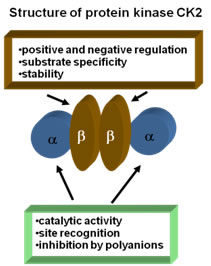
أثبت التحليل الكيميائي المناعي للنطاف وجود البروتين كيناز CK2 لديها، حيث استخدمت خمسة
فئران من مختلف الأعمار منها كشاهد، وذلك من مركز حيوانات التجربة التابع للمجموعة
البحثية للبروفسور ماتياس مونتينار في مدينة هومبرغ سار في ألمانيا وجرى العمل في مختبر
البيولوجيا الجزيئية والكيمياء الطبية، وذلك خلال صيف عام 2009.
وضعت الفئران ضمن دورة 12 ساعة ضوء و 12 ساعة ظلام مع الماء والطعام،
وذلك من أجل تنبيه الجهاز العصبي المركزي لديها، ثم جرى قتل الفئران من قبل المجموعة
المتخصصة والمدربة، في مركز حيوانات التجربة المذكور، باستخدام الخلع الرقبي cervical dislocation. كما جرى تطبيق التحاليل
الموصى بها من قبل تطوير المرشد لأجل رعاية واستخدام حيوانات التجربة في التعليم
والبحث العلمي.
عزل النطاف والكيمياء
الخلوية المناعية
قامت المجموعة المتخصصة
بالتعامل مع الحيوانات حسب الإرشادات القانونية المتبعة والتابعة للمجموعة البحثية
للبروفسور مونتينار في مركز البيولوجيا الجزيئية في ألمانيا، بعزل النطاف من
الفئران، وذلك بغمر بربخ الفأر في وسط wittingham medium دون ألبومين مصل البقر BSA ثم
تقسيمه إلى أجزاء صغيرة رفيعة باستخدام مشرط. تم حضن القطع بالدرجة 37م وضغط %5 CO2 في مستنبت chamber للسماح للنطاف بالسباحة خارج لمعة البربخ. نبذ المعلق بسرعة 3000 g لمدة 5 دقائق ثم غسلت الرسابة باستخدام
دارئة فسفاتية لإزالة الوسط العالق. استخدمت الرسابة لاستخلاص البروتين. لإنجاز
تحاليل الكيمياء الخلوية المناعية، علقت الرسابة في بارافورمألدهيد %4 لمدة ساعة.
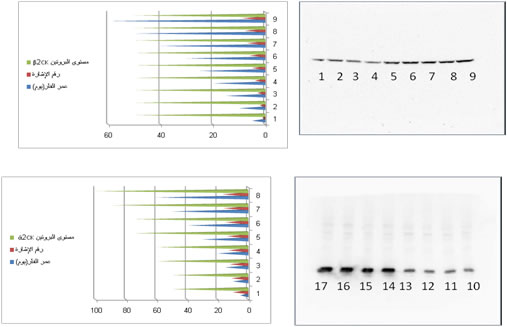
استخلاص البروتين ولطخة ويسترن western blot
جرى استخلاص البروتين بمجانسة النطاف المعزولة
كما ذكر سابقاً، حيث وضعت في الدارئة:
Buffer A (1% Triton X-100, 1 M NACL, 1 m M EDTA, 10 mg/ml phenylmethane
sulfonyl-fluoride, 20 mM TRIS-HCL PH 7, protease inhibitors leupeptin,
aprotinin and E-64).
ثم نبذت العينات لمدة 10 دقائق بسرعة 3000 g بدرجة 4م. حدد تركيز البروتين في السائل الطافي حسب
طريقة Bradford مع استخدام محلول BSA كمعياري (Bradford 1976). ومزج حوالي 100 µg من البروتينات مع دارئة lumli
3x، سخن الناتج لمدة 5 دقائق بالدرجة 95م من أجل أن تسمخ البروتينات. رحلت الخلاصات
الناتجة على هلامة عديد الأكريلاميد %12.5 SDS-polyacrylamide ثم نقلت إلى أغشية نتروسلولوز.
طبقت بعد ذلك تقانة western blot فحضنت الأغشية لمدة
ساعة مع دارئة الإحصار %2 blocking buffer، BSA/PBS وحضنت طوال الليل بدرجة 4م مع أضداد أولية نوعية ذوابة في محلول الإحصار وهي أضداد الوحيدة CK2ά متعددة النسائل مشتقة من البقر وبتركيز (400 ng/ml)، بالإضافة
إلى أضداد الوحيدة CK2β متعددة النسائل مشتقة من الأرانب وبتركيز (200 ng/ml)، وكلاهما من Santa Cruz Biotechnology, Calif;
USA أو β-actin (1 µg/ml;
Sigma, USA).
بعد ثلاث مرات غسل في الدارئة التالية: 0.1% Tween-20 in PBS (PBST) لمدة 10 دقائق في كل مرة، جرى كشف الأضداد الأولية باستخدام أضداد ثانوية
مقرونة بالإنزيم alkaline-phosphatase باستخدام نظام NBT/BCIP،
ثم أظهرت النتيجة بتطبيق نظام ECL. أعطى التفاعل مع الضد anti- CK2β عصابة واحدة فقط مع الأضداد المأشوبة his6-tagged
recombinant protein.
ولقد
برهن الامتزاز البدئي لأضداد anti- CK2ά مع ببتيد الإحصار الشاهد (Santa Cruz Biotechnology) أو مع CK2β المأشوب recombinant للضد العصابة الملاحظة في الشاهد.
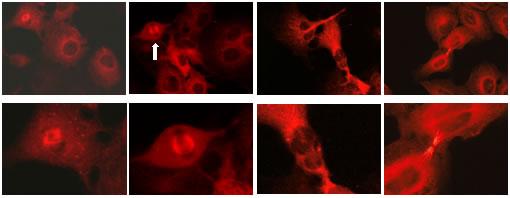
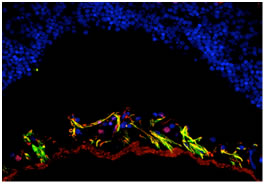
التألق المناعي Immunoflourescence
جرت مقايسة CK2 في نطاف الفأر المغمورة
بالبارافين 5 µm والمثبتة بمحلول Bouin. عوملت العينات بالماء
الأكسجيني 3% H2O2 مدة 5 دقائق، ولمنع الارتباط
غير النوعي طبق نظام إحصار البروتين المعياري لمدة 5 دقائق. جرى الغسل مرتين
لمدة 5 دقائق في دارئة TRIS-HCL بدرجة باهاء pH 7.6 مع 0.3 M NaCl و 0.1% Tween 20 ثم أذيبت الأضداد: أضداد الوحيدة CK2ά متعددة النسائل مشتقة من البقر (400 ng/ml) وأضداد الوحيدة CK2β متعددة النسائل مشتقة من
الأرانب (200 ng/ml)، كلاهما
من Santa Cruz Biotechnology, Calif; USA في محلول الإحصار ثم طبقت على العينات التي جرى حضنها طوال الليل
بدرجة 4م
في غرفة رطبة. بعد ذلك، أضيفت الأضداد التالية وجرى الحضن لمدة 15 دقيقة:
Biotinylated secondary antibody,
streptavidine-biotinlated-peroxidase complex, amplification reagent (biotinyltyramide),
and peroxidase-conjugated streptavidine (Lab-Vision)
أخيراً، أضيف محلول تلوين
الركازة (محلولDABI )
لمدة 5 دقائق وهو يتألف من:
Concentrated TRIS-HCL, 0.8% H2O2,
3,3-diaminobenzidine tetrahydrochloride (DABI)
غسلت الأجزاء بالماء المقطر. قرئت العينات
باستخدام مجهر تألق لشركة zies وصورت النتائج بكاميرا ديجيتال
باستخدام برنامج التألق المناسب على الحاسوب.
|