| المجلد 6 ,
العدد 2
, رجب 1432 - تموز (يوليو) 2011 |
| |
| محاولة التمنيع باستخدام اللَيشمانِيَّة مُثبّطة الانقسام بـ Mitomycin-c |
| Immunization Trial by Leishmania Treated with Mitomycin-c |
| ص. داليا العموري، أ.د. سعاد العقلة*، أ.د. محمود قويدر*، وأ.د. محمد معروف |
| Alammoury D; Kweider M;* Alokla S.* and Maarouf M. |
كلية الصيدلة، *كلية العلوم، جامعة دمشق
Faculty of pharmacy, *Faculty of science, Damascus University |
| الملخص Abstract |
| الليشمانية المدارية هي طفيلي من شعبة الأوالي تسبب مرضاً جلدياً في العالم القديم. هدفت الدراسة إلى تعيين المَناعَةٌ المُتَواسَطَةٌ بالخَلاَيا ( مَناعَةٌ خَلَوِيَّة) بقياس مستويات IFN-γ و IL-4 المصلية، في فئران BALB/C الممنعة بـ 107 من طفيليات الليشمانية الحية المعالجة بالميتوميسين-c (الليشمانية مثبطة الانقسام) (inhL)، الممزوجة مع 2 مكغ من IL-12 أوالممنعة بـ 2 مكغ من IL-12 فقط (الشواهد) أوالممنعة بـ 107 من طفيليات الليشمانية ذات النمط الشائع (المحتفظة بفوعتها) Wild type (WT). وجدنا أن inhL تحرض بشكل معتدٍ به مستويات أعلى من IFN-γ بمقدار 360.62 بكغ/ مل مقارنة مع الشواهد 4.36 بكغ/ مل ومع المجموعة الملقحة بـ WT 41.72 بكغ/ مل. وإلى جانب هذا تحرض inhL مستويات IL-4 أخفض 3.18 بكغ/ مل من الشواهد (أقل من 2 بكغ/ مل) ومن المجموعة المحقونة بـ WT حيث بلغ متوسط القيم 69.82 بكغ/ مل. أظهرت دراستنا أن inhL يحرض المَناعَةٌ المُتَواسَطَةٌ بالخَلاَيا مقارنة مع WT والشواهد. |
| Leishmania tropica is a protozoan parasite that causes cutaneous disease in the old world. The study aimed to determine cell-mediated immunity by measuring serum IFN-γ and IL-4 levels, in BALB/C mice immunized with 107 mitomycin-c treated live leishmania promastigotes (inhl), combined with 2µg IL-12, or immunized with 2µg IL-12 only(controls), or immunized with 107 wild type leishmania promastigotes. We found that inhl induce significantly higher levels of IFN-γ 360.62 pg/ml than controls 4.36 pg/ml and then WT 41.72 pg/ml. beside of this, inhl induce significantly lower IL-4 levels 3.18 pg/ml than WT 69.82 pg/ml and then controls (lower than 2pg/ml). Our study revealed that inhl induce cellular mediated immunity comparable with controls and WT. |
| المقدمة Introduction |
| داء الليشـمانية هو مجموعة من الآفات التـي تصيب الجلد والأغشية المخاطية والأحشاء، تسببه طفيليات أوالي من جنس الليشمانية Leishmania (1). ينتشر داء الليشمانية في 88 دولة موزعة بين آسيا وأوروبا وأفريقيا وأمريكا (2)، وتشمل هذه الدول سورية حيث تنتشر الإصابات الجلدية والحشوية بالليشمانية في عدة محافظات مثل دمشق وريفها، حلب، إدلب، الرقة، حمص، حماة والساحل السوري (3). يعاني المهتمون بمكافحة داء الليشمانية في المناطق التي يستوطن فيها، من صعوبة السيطرة على مخازن الطفيلي وصعوبة القضاء على الحشرة الناقلة ويضاف إلى ما سبق السمية الناجمة عن المعالجة بالأدوية الكيميائية وظهور الإصابة عند المثبطين مناعياً (4). وبالتالي تكتسب الوقاية من الإصابة بهذا الداء أهمية كبيرة وتتطلب وجود لقاح فعال مازال غير متوفر حتى الآن. نشرت العديد من الأبحاث حول لقاحات مختلفة تمت دراستها لكن لم يتوصل الباحثون، حتى الآن، إلى لقاح يمتاز بفعاليته ضد أنواع طفيليات الليشمانية جميعها. وبالتالي يكتسب العمل من أجل الوصول إلى لقاح فعال أهمية كبيرة في مجال الوقاية من الإصابة بهذه الطفيليات. هذا وتفضل اللقاحات المكونة من طفيليات حية على اللقاحات المكونة من البروتينات المؤشبة أو من خلاصة الطفيلي المقتول كاملاً، لأنها تبقى في الأنسجة، مما يؤمن ويحقق تزويداً مستمراً بالمستضد يدوم لفترة طويلة وبالتالي يحرض مناعة تستمر لفترة طويلة الأمد (5). ولهذا هدفت دراستنا إلى المساهمة في المحاولات الهادفة للوصول إلى لقاح فعال ولقد قررنا في سبيل ذلك استقصاء إمكان استخدام طفيليات حية مثبطة الانقسام للوصول إلى لقاح فعال ضد هذه الطفيليات. استخدمنا الميتوميسين Mitomycin-c، الذي يثبط تضاعف DNA الأشكال المتحركة لطفيليات الليشمانية (6)، لتثبيط انقسام سلالة الليشمانية المحلية المأخوذة من بلدنا. كما تحرينا قدرة هذه الطفيليات المثبطة على توليد المناعة الفعالة من خلال القيام بدراسة الاستجابة المناعية الخلوية، بمقايسة كل من IFN-γ و IL-4، عند حقنها للفئران من نمط BALB/C. وجرت مقارنة الاستجابة المناعية التي حرضتها هذه الطفيليات مع الاستجابة المناعية التي حرضتها الطفيليات المفوعة. ولقد بينت نتائجنا قدرة هذه الطفيليات المثبطة الانقسام على توليد استجابة مناعية خلوية جيدة وكانت من نمط Th1. |
| المواد والطرق Materials and Methods |
جرت هذه الدراسة في كل من كلية الصيدلة، جامعة دمشق، وكلية العلوم، جامعة دمشق، خلال الفترة الممتدة بين شهر أيلول 2008 إلى شهر تشرين الأول .2009
الطفيليات المستخدمة: جرى عزل سلالة الطفيليات المستخدمة في دراستنا من شاب من ريف دمشق، وبين تنميطها أنها من نمط الليشمانية المدارية L. tropica. استخدمنا وسط NNN لعزل الطفيليات واستخدمنا وسط RPMI 1640 لتكثير الطفيليات والمحافظة عليها أثناء الدراسة.
تحديد جرعة الميتوميسين-c الموقفة لتضاعف الأشكال المتحركة من الطفيليات: حُضّر محلول Mitomycin-c (Sigma) بتركيز 1000 مكغ/ مل. ثم قمنا بمعالجة الأشكال المتحركة من الطفيليات بتراكيز متزايدة من Mitomycin-c . وبعد ذلك حُضنت المزارع السابقة مدة 24 ساعة بدرجة حرارة 26°م، تم بعدها عد الطفيليات باستخدام عدادة نيوبار، مما سمح بتحديد الجرعة التي تثبط تكاثر الطفيليات خلال 24 ساعة.
تحضير اللقاح: حُضّر لقاح طفيليات الليشمانية المثبطة بالميتوميسسن- c بحضن 107 من الطفيليات المأخوذة من طور الاستتباب مع الميتوميسين–c بتركيز 12.7 مكغ مدة 24 ساعة. وجرى تحضير الطفيليات المفوعة بأخذ 107 من الأشكال المتحركة للطفيليات، في طور الاستتباب، وتعليقها بالمصل الفيزيولوجي. جرى الحصول على الإنترلوكين IL-12 الفأري المأشوب من شركة Sigma وجرى حله بالمصل الفيزيولوجي بتركيز 1000 مكغ/ مل. أُضيف 2 مكغ من المحلول السابق إلى الطفيليات المعالجة بالميتوميسين–c قبل حقنها للفئران.
الفئران المستخدمة: استخدمنا فئران BALB/C، بلغت أعمارها 6 أسابيع. قسمت هذه الفئران إلى ثلاث مجموعات تحوي كل منها خمسة فئران. حُقنت هذه الفئران في أسفل القدم تحت الجلد. حيث حقنت فئران المجموعة الأولى بـ 107 من الأشكال المتحركة للطفيلي المعالجة بالميتوميسين–c والممزوجة مع 2 مكغ من IL-12. وحقنت فئران المجموعة الشاهدة Control بالمصل الفيزيولوجي الحاوي على 2 مكغ rm IL-12 . وحقنت فئران المجموعة الثالثة بـ 107 من الطفيليات المفوعة.
الإعتيان :Sampling جرى بزل الدم من الفئران بعد مرور 4 أسابيع على عدوى الفئران بالطفيليات وذلك لمقايسة الإنترفيرون غاما والإنترلوكين 4-.
جرت مقايسة الإنترفيرون-غاما في مصول الفئران باستخدام عتيدة مُقَايَسَةُ المُمْتَزِّ المَناعِيِّ المُرْتَبِطِ بالإِنْزِيْم (ELISA) من شركة MABTECH لمقايسة الإنترفيرون الفأري Mouse IFN-γ. وجرت مقايسة الإنترلوكين IL-4 في مصول الفئران باستخدام عتيدة Mouse IL-4 ELISA لشركة BioVendor.
|
| النتائج Results |
-1 بينت نتائجنا أن الميتوميسين–c يثبط انقسام الطفيليات المدروسة، وتتناسب شدة التثبيط طرداً مع ارتفاع تراكيز الميتوميسين المتزايدة (كما يبين الشكل (1. ومن خلال معادلة المنحنى البياني، Y=0.668x + 18.1، للعلاقة بين التأثير المثبط للتراكيز المختلفة من الميتوميسين–c وأعداد هذه الطفيليات، قمنا بحساب الجرعة المثبطة لتكاثر 107 من الأشكال المتحركة للطفيلي والتي بلغت 12.7 مكغ/ مل.
-2 قيم الإنترفيرون غاما: يلخص الجدول 1 المتوسطات الحسابية والانحرافات المعيارية لقيم الإنترفيرون غاما لكل مجموعة من المجموعات. حيث بلغ تركيز IFN-γ 4.36 بكغ/ مل لدى المجوعة الشاهدة و 41.72 بكغ/ مل لدى المجموعة المحقونة بـ 107 من الطفيليات المفوعة، و 360.62 بكغ/ مل في مصول الفئران المحقونة بـ 107 من الليشمانية الحية المثبطة بالميتوميسين.
ونستطيع المقارنة من خلال الشكل 2 بين النتائج السابقة، والتي تظهر ارتفاع قيم IFN-γ لدى المجموعة المحقونة بالليشمانية الحية المثبطة الانقسام بالميتوميسين inhL مقارنة مع المجموعة الشاهدة وبفارق يعتد به إحصائياً P<0.001. كما لوحظ ارتفاع في قيم IFN-γ لدى المجموعة المحقونة بالليشمانية المفوعة مقارنة مع المجموعة الشاهدة بفارق يعتد به إحصائياً P<0.01.
كما ترتفع قيم IFN-γ لدى المجموعة المحقونة بالليشمانية الحية المثبطة الانقسام بالميتوميسين مقارنة مع المجموعة الملقحة بالليشمانية المفوعة وكان الفارق ذا دلالة إحصائية (P<0.001).
3- قيم الإنترلوكين-4: يلخص الجدول 2 المتوسطات الحسابية والانحرافات المعيارية لقيم الإنترلوكين – 4 لكل مجموعة من المجموعات. حيث بلغ تركيز IL-4 69.82 بكغ/ مل لدى المجموعة المحقونة بـ 107 من الليشمانية المفوعة. وبلغ 3.18 بكغ/ مل في مصول الفئران المحقونة بـ107 من الليشمانية الحية المعالجة بالميتوميسين، ولكنه كان أقل من 2 بكغ/ مل، حيث تشكل 2 بكغ/ مل عتبة الحساسية بالنسبة للعتيدة المستخدمة، في مصول الفئران الشاهدة.
ونستطيع المقارنة من خلال الشكل 2 بين النتائج السابقة، والتي تظهر ارتفاع قيم IL-4 لدى المجموعة المحقونة بالليشمانية الحية المثبطة الانقسام بالميتوميسين مقارنة مع المجموعة الشاهدة وبفارق يعتد به إحصائياً، حيث بلغت قيمة P<0.01 . كما لوحظ ارتفاع في قيم IL-4 لدى المجموعة المحقونة بالليشمانية المفوعة مقارنة مع المجموعة الشاهدة بفارق يعتد به إحصائياً (P<0.001). ولكن تنخفض قيم IL-4 لدى المجموعة المحقونة بالليشمانية الحية المثبطة الانقسام بالميتوميسين مقارنة مع المجموعة المحقونة بالليشمانية المفوعة وبفارق يعتد به إحصائياً (P<0.001).
|
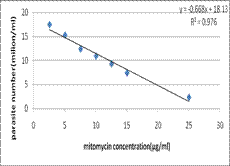
الشكل 1: العلاقة بين تراكيز مختلفة للميتوميسين وعدد الطفيليات بعد حضن 10 مليون طفيلي لمدة 24 ساعة.
الجدول 1: المتوسط الحسابي والانحراف المعياري لتراكيز IFN-γ لدى مجموعات الدراسة.
|
تركيز الإنترفيرون γ بيكوغرام/ مل |
المجموعة الشاهدة |
الطفيليات المفوعة |
الطفيليات المثبطة |
X |
SD |
X |
SD |
X |
SD |
الفأر1 |
3.5 |
0.56 |
43.5 |
3.25 |
374.55 |
8.13 |
الفأر2 |
6.1 |
0.28 |
62.2 |
2.96 |
421.3 |
13.85 |
الفأر3 |
5.11 |
0.43 |
54.5 |
3.81 |
321.6 |
8.34 |
الفأر4 |
2.8 |
0.42 |
29.5 |
3.95 |
299.76 |
2.19 |
الفأر5 |
4.3 |
0.28 |
18.9 |
2.26 |
385.9 |
6.78 |
المتوسط |
4.36 |
1.16 |
41.72 |
17.72 |
360.62 |
49.3 |
الجدول 2 : المتوسط الحسابي والانحراف المعياري لتراكيز IL-4 لدى مجموعات الدراسة.
|
تركيز IL-4 (بيكوغرام/ مل) |
المجموعة الشاهدة |
الطفيليات المفوعة |
الطفيليات المثبطة |
X |
SD |
X |
SD |
الفأر1 |
أقل من 2 |
52.98 |
1.69 |
2.1 |
2.28 |
الفأر2 |
أقل من 2 |
78.65 |
0.64 |
3.48 |
0.86 |
الفأر3 |
أقل من 2 |
80.93 |
1.68 |
4.76 |
0.86 |
الفأر4 |
أقل من 2 |
79.11 |
0.45 |
2.76 |
0.36 |
الفأر5 |
أقل من 2 |
57.44 |
0.22 |
2.44 |
0.29 |
المتوسط |
أقل من 2 |
69.82 |
13.45 |
3.18 |
1.09 |
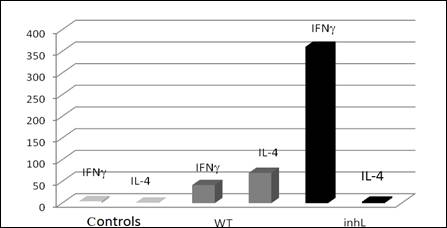
الشكل 2 : متوسط قيم الإنترفيرون غاما والإنترلوكين 4 لدى مجموعات الدراسة. 5 فئران في كل مجموعة. أُجريت تجربتان لكل فأر، حيث inhL الليشمانية الحية المثبطة الانقسام بالميتوميسين، WT الطفيليات المفوعة، Controls الشواهد. |
| المناقشة Discussion |
| إن تحديد نمط الاستجابة المناعية Th1 أو Th2 هو الذي يحدد فعالية اللقاح. حيث تعد الاستجابة المناعية المتواسطة بالخلايا المتميزة بإفراز IFNγ 6))، الذي يقوم بتحريض البلاعم على تشكيل الجذور النترية الحرة، الأساسية في القضاء على طفيلي الليشمانية داخل الخلوي (7). بينما تدخل الاستجابة المناعية من نمط Th2 المتميزة بإفراز IL-4 لمنع التَّأثيرات الجيدة لـ IFN-γ (8، 9). لا تزال الآليات المسؤولة عن تمايز Th1 عند الفئران المقاومة والآليات المسؤولة عن تمايز Th2 عند الفئران الحساسة مثل الفئران BALB/C غير مفهومة بشكل كامل. هذا ويعتقد بأنَّ الآليات السابقة تعود إلى الفروق في السيتوكينات المفرزة باكراً لحظة تماس الخلايا التائية غير المتمرسة مع المستضد. ويعد IL-12 من أهم هذه السيتوكينات المفرزة في المراحل الباكرة من الإصابة (10). وهذا ما قادنا إلى استخدام IL-12 كعامل مساعد في دراستنا هذه.
بينت نتائجنا أن الميتوميسين-c يثبط انقسام الطفيليات المدروسة وهذا التركيز يتوافق مع تركيز الميتوميسين-c المستخدم في تثبيط تكاثر الليشمانية في دراسة Reiner E. (11).
أظهرت دراستنا ارتفاع قيم IFN-γ في مصول الفئران المحقونة بمزيج من الطفيليات الحية المعالجة بالميتوميسين–c و IL-12 مقارنة مع القيم المقابلة لها لدى مجموعة الفئران المحقونة بـ IL-12 فقط وكان هذا الفارق معتداً به إحصائياً. كما سجلنا ارتفاعاً بسيطاً جداً في قيم IL-4 في مصول الفئران المحقونة بمزيج من الطفيليات الحية المعالجة بالميتوميسين-c وIL-12 مقارنة مع القيم المقابلة لها لدى مجموعة الفئران المحقونة بـ IL-12 فقط. وبالتالي سجلنا ارتفاعاً في نسبة الإنترفيرون غاما إلى IL-4، التي بلغت 113.4، لدى المجموعة المحقونة بمزيج من الطفيليات الحية المعالجة بالميتوميسين–c وIL-12 مقارنة مع قيمة النسبة المقابلة لها لدى مجموعة الفئران المحقونة بــ IL-12 فقط والتي بلغت 2.16. ويمكن أن يعود ارتفاع النسبة السابقة إلى أن الطفيلي يقوم بدور محفز للمناعة وأن IL-12 يوجه الاستجابة المناعية نحو نمط Th1 في مجموعة الفئران المحقونة بالطفيليات الحية المثبطة بالميتوميسين –c و IL-12. كما بينت نتائجنا ارتفاع قيم IFN-γ وانخفاض قيم IL-4 مقارنة مع الفئران المحقونة بـالطفيلي المحتفظ بفوعته، ويمكن أن يعود ذلك إلى وجود مستضدات في الليشمانية مثل مستضد LACK ومستضدLmI1 تقوم بخداع الجهاز المناعي وإجباره على تنشيط الخلايا Th2 (12). كما تؤدي المستضدات السابقة إلى تحريض Th1 المنتجة لـ IFN-γ، إذا تشاركت مع IL-12 (13) الذي استخدمناه كمادة مساعدة في مجموعة الفئران المحقونة بالطفيليات الحية المعالجة بالميتوميسين-c. ونظراً لارتفاع قيم IFN-γ وانخفاض قيم IL-4 في مصول الفئران المحقونة بالطفيليات الحية المثبطة بالميتوميسين–c وIL-12 بالمقارنة مع كل من المجموعة الشاهدة والمجموعة الملقحة بالطفيليات المفوعة، فإننا نرجح وجود استجابة مناعية خلوية من نمط Th1 لدى فئران المجموعة المحقونة بالطفيليات الحية المثبطة الانقسام. أما بالنسبة لموضوع استخدام طفيليات مثبطة الانقسام بالميتوميسين فلم تستخدم هذه الطفيليات مسبقاً، لكن هنالك عدة دراسات استخدمت طفيليات معدلة جينياً (13-17) ولقد توافقت نتائجنا مع نتائج تلك الدراسات التي حرضت فيها الطفيليات السابقة استجابة مناعية من النمط Th1.
أما بالنسبة لمجموعة الفئران المحقونة بالطفيليات المفوعة بدون مشاركة IL-12، فلقد بينت نتائجنا ارتفاع قيم IL-4 و IFN-γ مقارنة مع مجموعة الفئران المحقونة بـ IL-12 فقط. و يشير ذلك إلى وجود استجابة مناعية مختلطة تغلب عليها استجابة Th2 بسبب القيم المرتفعة من IL-4. تتوافق هذه النتيجة مع اختيار نوع الفئران BALB/C، وهي فئران حساسة للإصابة بالليشمانية (18)، حقنت بالطفيليات المفوعة وبدون مشاركة أي مادة مساعدة، وبالتالي يعد ظهور استجابة مناعية من النمط Th2 أمراً طبيعياً في هذه الحالة.
|
| الاستنتاج Conclusion |
|
حرضت الليشمانية المدارية المثبطة الانقسام، بمشاركة IL-12 استجابة مناعية خلوية من نمط Th1 عند الفئران الحساسة BALB/C، وهذا يعد عاملاً إيجابياً في ما يخص إمكان استخدام الطفيليات السابقة كلقاح.
|
| المراجع References |
1-Dejeux P.
Leishmaniasis: current situation and new perspectives.
Comp. Immaunal. Microvial. Intect. Dis. 27(5): 18-305, 2004.
2-Klaus SN; Frankenburg S. and Ingber A.
Epidemiology of cutaneous leishmaniasis.
Clin Dermatol. 17(3): 257-260, 1999.
3-Marouf M.
Distribution of cutaneous lishmaniasis in Damascus and its suburbs during 2002-2005.
Arab Journal of Pharmaceutical Sciences, 3(3): 39-51, 2006.
4-Clarisa B. and Palatnik-de-Sousa
Vaccines for leishmaniasis in the fore coming 25 years .
Vaccine. 26, 1709-1724, 2008.
5-PST Veras, CI Brodskyn FMP Balestieri and LAR de Freitas
A dhfr-ts- Leishmania major Knockout Mutant Cross-protects against Leishmania amazonensis.
Mem Inst Oswaldo Cruz, Rio de Janeiro, 94(4): 491-496, 1999.
6-Chahnaz Kébaïer, Jude E. Uzonna, Stephen M. Beverley and Phillip Scott.
Immunization with Persistent Attenuated ?lpg2 Leishmania major Parasites Requires Adjuvant To Provide Protective Immunity in C57BL/6 Mice.
Infect Immun. 74(1): 777-780, 2006.
7-Alexander J; Graham H. Coombs and Jeremy C. Mottram.
Leishmania mexicana Cysteine Proteinase-Deficient Mutants Have Attenuated Virulence for Mice and Potentiate a Th1 Response1.
Immunology. 161, 6794-6801,1999.
8-Saravia N.G; Escorcia B, Osorio Y. et al.
Pathogenicity and protective immunogenicity of cysteineproteinase-deficient mutants of Leishmania mexicanain non-murine models.
Vaccine. 24, 4247-4259, 2006.
9-Handman E.
Leishmaniasis: Current Status of Vaccine Development .
Clinical microbiology Reviews. 14(2): 229-243, 2001.
10-Reiner E. and Kazura W.
oxidant-mediated damage of leishmania dononvani promastigote.
Infection and immunity. 36(3): 1023-1027, 1999.
11-Mougneau E; Altare F. et al.
Expression cloning of a protective Leishmania antigen.
Science. 58, 2198-2203, 1995 .
12-Audrey Y. Park, Brian D. Hondowicz and Phillip Scott.
IL-12 Is Required to Maintain a Th1 Response During Leishmania major Infection.
Immunology. 165, 896-902, 2000.
13-Roberts M.T.M.
Current understandings on the immunology of leishmaniasis and recent developments in prevention and treatment.
British Medical Bulletin. 44, 75-76, 2006.
|
| |
| المجلد 6 ,
العدد 2
, رجب 1432 - تموز (يوليو) 2011 |
|
|
|
